



Inscriete la noutatile noastre
Livrăm gratuit în raza municipiului Chișinău comenzile ce depășesc valoarea de 500 lei.
31 Mar 2022
Principalele boli la suine
Pesta porcină clasică (PPC)
Etiologie
Virusul pestei porcine clasice aparține genului Pestivirus din familia Flavivirdae, este un virus ARN monocatenar, de dimen- siuni mici (40-60nm), formă sferică și prezintă capsidă. Posibilitatea de recombinare între tulpinile sale este foarte mare.
Epidemiologie
Este o boală infecțioasă foarte contagioasă, cu importanță la nivel mondial. Populațiile de porci din SUA, Australia, Noua Zeelandă și din majoritatea țărilor UE sunt declarate indemne. PPC a rămas endemică în Africa și Asia. Calea principală de transmitere este cea oro-nazală, prin contact
direct sau indirect cu animale infectate sau prin ingestie de furaje contaminate. Nerespectarea normelor de biosecuritate în ferme poate avea un rol major în transmiterea acestui virus. Animalele infectate elimină virusul prin secreții și excreții.
Patogeneză
Replicarea inițială a virusului se realizează la nivelul tonsilelor, ulterior difuzând la țesuturile limfoide din zonă. De aici, se răspândește la nivelul limfonodurilor regionale pe cale limfatică. Aici are loc din nou replicarea, iar virusul este răspândit prin intermediul sângelui la locurile de replicare
secundară precum: măduva hematogenă, splina, măduva osoasă, limfonoduri și țesuturi limfoide asociate viscerelor. Răspândirea virusului în întreg organismul are loc în mai puțin de 6 zile. Replicarea virusului are loc atât în monocite, macrofage, cât și în celulele endoteliale.
Apoptoza precum și activarea fagocitică și secretorie pot fi observate în populațiile de macrofage. Macrofagele activate par să joace un rol important în patogeneza infecției cu virusul PPC, în timp ce leziunile cauzate direct de către virus pot fi excluse aproape în totalitate. În forma
acută, letală, PPC este însoțită de limfopenie severă, imunosupresie consecutivă și granulocitopenie; mai mult, imediat după infecție poate fi observată o trombocitopenie severă. Mecanismele trombocitopeniei nu sunt pe deplin înțelese, dar sunt implicate: coagularea intravasculară
diseminată (CID), leziuni ale măduvei hematogene și degenerarea megacariocitelor. Mecanismul patogen implicat în apariția leziunilor hemoragice cuprinde injurii ale celulelor endoteliale, trombocitopenia, CID, eritrodiapedeza, vasodilatația capilară și permeabilitatea crescută.
Tablou clinic
În forma acută, letală, moartea indivizilor apare la 2-4 săptămâni după infecția cu virusul PPC. Mortalitatea poate să ajungă la 100% între zilele 10 și 30, în funcție de vârsta animalului și de virulența tulpinii virale. Din cauza caracterului imunosupresiv al PPC, infecțiile secundare
severe ale sistemului respirator și gastrointestinal pot complica evoluția bolii, putându-se suprapune cu semnele clinice ale PPC. Semnele clinice inițiale includ anorexie, letargie, conjunctivită, semne respiratorii și diaree urmată de constipație. Forma cronică evoluează atunci
când un individ infectat nu este capabil să genereze un răspuns imun adecvat. În general, animalele infectate prezintă semne clinice nespecifice, precum febră intermitentă, epuizare sistemică, enterită cronică și dermatită difuză. S-a demonstrat faptul că toate animalele infectate
cu forma cronică vor muri în cele din urmă, dar pot supraviețui câteva luni de la infecție, timp în care pot elimina, constant, cantități impresionante de virus. Când infecția apare la scroafele gestante, virusul poate infecta fătul, datorită capacității sale de a traversa bariera
placentară, ceea ce poate duce la infecții persistente la purcei. În timp ce scroafele prezintă adesea doar semne clinice ușoare, această infecție poate duce, în funcție de stadiul gestației, la absorbția sau mumificarea fetușilor, la avorturi sau la nașterea de purcei neviabili. Atunci când infecția se
produce între zilele 50 și 70 ale gestației, apare un fenomen de imunotoleranță, iar purceii vor suferi o infecție persistentă.Inițial purceii par sănătoși și supraviețuiesc câteva luni, dar mor apoi din cauza unei forme a PPC cu debut tardiv. În această perioadă purceii elimină virus în cantități
mari, suficiente pentru transmitere.
Leziuni anatomopatologice În forma supraacută apare eritem cutanat, peteșii la nivelul seroaselor, mucoaselor, precum și în cortexul renal.
Limfonodurile sunt congestionate. În forma acută predomină un tablou hemoragic însoțit de leucopenie, trombocitopenie, peteșii și echimoze la nivel cutanat, precum și la nivelul limfonodurilor, laringelui, vezicii urinare, rinichilor și intestinului subțire. Infarctele marginale splenice,
multiple, sunt caracteristice în PPC, dar nu sunt întotdeauna prezente. Limfonodurile și tonsilele apar edematoase și hemoragice. Tot în forma acută se observă conjunctivite sero-purulente, eritem și infarcte cutanate, diateză hemoragică, hemoragii limfonodale (limfonodurile prezentate cu un
aspect marmorat) și infarcte multifocale la nivel splenic. Pe fond distrofic apar multiple peteșii la nivelul corticalei renale, bazinetului renal și vezicii urinare. Se observă necroza multifocală a tonsilelor, hemoragii laringiene și gastrice (sub seroase). Leziunile de colită hemoragico-necrotică sunt
deseori întâlnite.
În forma subacută se evidențiază angină ulcero-necrotică, gastrită hemoragico-catarală, necroza plăcilor limfoide însoțită de inflamații difteroide, tiflo-colită difteroidă în focare sub forma „butonilor pestoși“, hemoragii pulmonare,bronhopneumonii fibrino-necrotice, gangrenoase
și exantem varioloid.
.png)
În forma cronică apar butonii pestoși la nivelul cecumului și colonului. Leziunile hemoragice sau inflamatorii sunt mai puțin întâlnite în această formă. Se observă, de asemenea, atrofia timusului, colită difteroidă și ulceroasă multifocală, bronhopneumonie fibrino-necrotică, gangrenoasă, pleurită fibrinoasă și sfacelări cutanate. Purceii nou-născuți din scroafe bolnave prezintă hipoplazie sau aplazie cerebrală, cerebeloasă, artrogripoză; frecvent sunt expulzați fetuși mumifiați.
La examinarea histopatologică a limfonodurilor și tonsilelor se observă congestie și hemoragii multiple. Foliculii limfoizi sunt afectați de depleție, fiind observate detritusuri celulare și hemoragii pe fond de necroză. La nivel splenic se observă grade diferite de hemoragie, de la focală la difuză,
în pulpa roșie. Rinichii prezintă congestie, hemoragii focale și nefrită interstițială cu celule mononucleare.
La nivel pulmonar apare pneumonie interstițială cu hemoragii extensive și exsudat serofibrinos în spațiile alveolare. La nivel hepatic, hepatocitele din zona centrolobulară prezintă modificări degenerative cu focare de necroză. În plăcile
Peyer de la nivelul ileonului pot fi observate aspecte de depleție limfoidă. Apar leziuni de encefalomielită limfoglială cu hialinizarea pereților vasculari și tromboze.
Diagnosticul de laborator Supravegherea serologică se poate realiza prin tehnica ELISA, utilizând sânge recoltat pe heparină. Pentru diagnosticul de certitudine se utilizează tehnica PCR, hibridizarea in situ, imunohistochimia și imunofluorescența directă utilizând sânge integral, tonsile, splină, ileon și limfonoduri.
Pesta porcină africană (PPA)
Etiologie
Virusul pestei porcine africane este un virus cu ADN de formă icosaedrică și dimensiuni mari, fiind singurul membru al familiei Asfarviridae, genul Asfivirus.
Epidemiologie
Pesta porcină africană (PPA) a evoluat ca o boală endemică pe continentul african, urmând să se răspândească în Asia, Europa și America. Atât suinele domestice cât și cele sălbatice sunt gazde naturale pentru virus. Porcii mistreți europeni sunt la fel de susceptibili la infecție, prezintă semnele clinice și rata mortalității similare cu cele observate la porcii domestici. Speciile sălbatice de suine de pe continentul african nu prezintă semne clinice și au rol de rezervor. Toate fluidele și țesuturile porcilor domestici cu semne clinice conțin cantități mari de virus. Virusul rezistă câteva luni în carnea refrigerată și ani în cea congelată.
Patogeneză
Virusul pestei porcine africane se transmite între suine cel mai frecvent pe calea contactului oro-nazal, dar și prin consum de furaje contaminate sau pe cale iatrogenă. Replicarea primară se realizează la nivelul monocitelor și macrofagelor de la nivelul tonsilelor și a limfonodurilor capului și
gâtului. De aici, virusul se răspândește pe cale sanguină sau limfatică în locurile de replicare secundară: limfonoduri, măduvă osoasă, splină, plămâni, ficat și rinichi. Virusul se replică atât în monocite și macrofage, cât și în megacariocite, celule endoteliale, hepatocite, celule epiteliale tonsilare, renale și neutrofile. Virusul induce apoptoza în celulele infectate cât și în limfocitele neinfectate.
Se consideră că patogeneza virală și semnele clinice sunt produse, în mare parte, de citokinele proinflamatorii produse de monocite/macrofage și alte celule infectate.
Patogeneza hemoragiei este atribuită alterării endoteliului și trombocitopeniei care induc disfuncția mecanismelor coagulării.
Hemoragia observată în forma acută se datorează activării fagocitozei celulelor endoteliale în care a avut loc replicarea virusului, iar în forma subacută,se datorează în principal creșterii permeabilității vasculare. Infecția transplacentară cu tulpini cu virulență scăzută poate duce la infecția
persistentă a purceilor.
Tablou clinic
Perioada de incubație în infecțiile naturale variază între 4 și 19 zile. Procentul de morbiditate variază în funcție de tulpina virală, calea de expunere și de prezența sau absența leziunilor hemoragice. Formele clinice ale pestei porcine africane variază de la supraacute (moarte subită, fără sau cu semne clinice minore) până la infecție subclinică. În formele supraacute și acute, semnele clinice sunt dominate de anorexie, letargie, stare de slăbiciune, decubit, hipertermie (40-42ºC), leucopenie marcantă, eritem și cianoză a urechilor, a cozii, a membrelor
și abdomenului. Eritemul, hemo- ragiile cutanate, melena, epistaxisul, morbiditatea și mortalitatea mare sunt caracteristice.
Scroafele gestante avortează.
Moartea se produce în 1-3 zile în formele supraacute și în 7-10 zile în cazul formelor acute, în aproape 100% din cazuri. Porcii cu forma subacută a bolii prezintă leucopenie severă, dispnee, vomă, jetaj și epiforă. Unii indivizi prezintă hipertermie recurentă, încetinirea creșterii și emaciere.
Porcii mor sau își revin în 3-4 săptămâni. Forma cronică se caracterizează prin febră joasă, pierderea apetitului, îngroșarea articulațiilor, ocazional diaree și vomă, stare generală afectată și prezența ulcerelor cutanate.
Leziuni anatomopatologice
Formele supraacută și acută de boală sunt caracterizate de congestie, edem și hemoragii în diverse țesuturi. Se observă eritem și sufuziuni cutanate, iar la nivel subcutanat edem și hemoragii. Apar peteșii la nivelul seroaselor și, uneori, colecții de lichid seros sau serohemoragic
în marile cavități seroase. Edemul pulmonar, hidrotoraxul și hidropericardul apar frecvent. Sunt întâlnite hematoame musculare, perirenale și la nivelul bazinetul renal. Splina este mărită mult în volum, închisă la culoare, friabilă, cu hemoragii. Meningele poate fi congestionat, edematos
sau hemoragic. Hemoragiile pot fi întâlnite și la nivelul mucoaselor digestive.
În limfonoduri apar hemoragii difuze masive și necroză, încât limfonodurile par hematoame. Ficatul apare congestionat iar la nivelul mucoasei veziculei biliare pot fi observate multiple peteșii și edem. Apare edem perirenal, peteșii multiple la nivelul corticalei renale ce pot conflua,
precum și hemoragii medulare. În unele episoade au fost raportate hemoragii și necroze pancreatice extinse. Doar splenomegalia severă, edemul pulmonar sever și limfonodurile hemoragice asemănătoare hematoamelor sunt caracteristice PPA, celelalte modificări fiind întâlnite și în PPC.
În forma subacută hemoragiile sunt mai restrânse, apărând la nivelul mucoasei intestinale, în rinichi și limfonoduri. Splina este mărită, dar necongestionată și apar consolidări în lobii pulmonari anteriori.
Forma cronică se caracterizează prin hiperplazie limfoidă (mărirea în volum a splinei și a limfonodurilor), pleurită și pericardită fibrinoasă, pneumonie fibrinoasă ce poate deveni necrotică și cu leziuni distrofice de calcificare. Se mai pot întâlni leziuni cutanate, de la eritem până
la zone de necroză, precum și îngroșarea articulațiilor - artrite.
Fetușii avortați pot prezenta anasarcă, peteșii cutanate și miocardice, iar pulmonii și ficatul pot căpăta un aspect marmorat. Histopatologic, virusul PPA, spre deose- bire de cel PPC, produce apoptoza și necroza extensivă a macrofagelor și limfocitelor.
Alterarea endoteliului și trombocitopenia vor induce coagulare intravasculară diseminată, hemoragii și edeme. Consecutiv, la nivel pulmonar se produce îngroșarea pereților alveolari. Hemoragiile și necrozele țesuturilor limfoide sunt severe. Degenerarea epiteliului renal va duce la formarea ci-
lindrilor amorfi medulari. Necroza hepatocitelor periportale cu infiltrat limfocitar este o leziune frecventă. Perivasculita în sistemul nervos central este asociată cu multe celule aflate în necroză. La fel ca în cazul PPC, PPA induce alterarea endoteliului vascular și necroza fibrinoidă a vaselor de sânge.
Forma cronică se caracterizează prin hiperplazie limfoidă, pleurită și pericardită fibroasă, pneumonie, meningoencefalomielită, glomerulonefrită segmentală focală și hepatită periportală.
Diagnosticul de laborator Investigațiile de laborator sunt absolut necesare pentru confirmarea acestei boli, deoarece există similitudini între PPA și alte patologii hemoragice ale porcilor. Cea mai utilizată metodă de identificare a antigenului este PCR, dar se poate folosi cu succes
hibridizarea in situ și imunofluorescența directă. Probele recomandate sunt limfonodurile viscerale, splina și sângele.
Circoviroza porcină
Etiologie
Circovirusul porcin aparține genului Circovirus din familia Circoviridae, fiind un virus fără capsidă, cu un diametru cuprins între 12 și 23 nm. Acest virus este responsabil de apariția sindromului porcin de epuizare multisistemică post-înțărcare (engl. PMWS), a sindromului porcin dermatitănefrită
(engl. PDNS), cât și a unui sindrom porcin reproducător (engl. PRD).
Epidemiologie
PCV-1 este ubicvitar în populațiile de suine, dar prevalența sa este mult mai mică față de PCV-2, fiind mai întâlnit la suinele sălbatice. PCV-2 are o prevalență foarte mare atât la suinele domestice cât și la cele sălbatice, ambele fiind gazde naturale. Recent au fost introduși termenii
PCV2-SD, pentru boala sistemică, PCV2-RD pentru boala reproducătoare și PCV2-SI pentru forma unei infecții subclinice.
Transmiterea oro-nazală este considerată calea principală de transmitere. PCV a fost identificat în secreții nazale, amigdaliene, bronhice și oculare, fecale, salivă, urină, colostru, lapte și material seminal. Porcii se pot infecta și prin consumul de țesuturi crude de la animale infectate.
Transmiterea
PCV-2 între indivizi se realizează atunci când se introduc animale purtătoare într-un efectiv sănătos. Contactul direct între animale mărește considerabil șansele de transmitere.
Studiile longitudinale ce cuantifică PCV-2 din ser, secreții nazale și fecale au arătat faptul că vârsta medie de infecție este între 4-11 săptămâni, în funcție de exploatație. Indivizii care manifestă semne ale PCV2-SD prezintă co-infecții cu alți agenți biologici, sugerându-se faptul că o infecție
cu PCV-2 asociată cu prezența altor factori este necesară pentru evoluția clinică com- pletă. Nivele mari de PCV-2 în sânge, țesuturi limfatice și limfoide, precum și lichidele de excreție sunt asociate cu expresia clinică a bolii. PCV-2 poate persista în organism pentru câteva luni, ceea ce face ca indivizii în convalescență să fie purtători și eliminatori de virus pentru perioade extinse.
Patogeneză
Contaminarea se realizează prin fluide oro-nazo-faringiene, fecale și urină de la animalele infectate. Virusul inhalat sau in- gerat pătrunde în celule epiteliale, macrofage, celule dendritice, sau celule M pe la nivelul plăcilor Peyer. Mecanismul de injurie în PMWS este liza și disfuncția limfocitelor
induse viral, implicit depleția limfocitară și imunosupresie.
.png)
Pentru infecție și replicare, virusul are nevoie de celule gazdă aflate în curs de diviziune, precum limfocitele în faza S. Viremia se poate detecta la 7 zile și ajunge la un nivel maxim la 14-21 zile post infecție. Anumite linii genetice de suine
sunt mai susceptibile față de altele. Probabilitatea ca indivizii din rasa Landrace să dezvolte PMWS este mult mai mare decât la indivizii din rasele Duroc, Marele alb sau
Pietrain. PCV-2 este capabil de replicare atât la nivel fetal, cât și în blastociștii eclozați (cu zona pelucida resorbită), ceea ce poate duce la moarte embrionară. De asemenea, a fost demonstrată și transmiterea transplacentară. PDNS este considerată ca o reacție de hipersensibilitate de tip III, în care antigenul prezent în complexele imune este necunoscut.
Tablou clinic
a. Sindromul de epuizare multisistemică post-înțărcare (PMWS) Sindromul este caracterizat prin pierderea excesivă în greutate. Purceii cu vârste cuprinse între 2 și 4 luni sunt cei mai susceptibili să dezvolte PMWS. Morbiditatea este de 5% - 20%, iar mortalitatea poate trece ușor peste 50%. Pe lângă pierderile datorate mortalității, poate determina o prelungire substanțială a timpului necesar pentru a atinge greutatea cerută de piață, ceea ce duce la pierderi economice. Semnele clinice cuprind: retard în creștere, stare generală de epuizare, paloare cutanată, insuficiență respiratorie, diaree și uneori icter. În prima parte a fazei clinice a bolii, limfonodurile superficiale sunt reacționate.
Suprapopularea, calitatea slabă a aerului, ventilația improprie, amestecarea indivizilor din diferite categorii de vârstă pot exacerba evoluția bolii. Debutul patologiei poate fi unul brusc, ducând la moarte în doar câteva zile a unor indivizi. Alți indivizi sunt afectați de o formă mai cronică a bolii, manifestând retard în creștere. Principalele caracteristici anamnetice ale acestei patologii sunt: tremor congenital, avort, purcei mumifiați,
cuiburi neuniforme.
b. Sindromul dermatită-nefropatie (PDNS) Sindromul afectează purceii nou-născuți, pe cei în creștere cât și adulții. Mortalitatea se apropie de 100% la porcii cu vârste mai mari de 3 luni, iar la porcii mai tineri ajunge la aproximativ 50%. Porcii cu o evoluție acută, severă, mor în câteva
zile după debutul semnelor clinice datorită insuficienței renale acute, cu o creșter semnificativă a nivelelor serice ale creatininei și ureei. Indivizii care supraviețuiesc tind să se recupereze și să crească în greutate la 7-10 zile după încetarea sindromului. Indivizii afectați prezintă anorexie
și abatere, fără sau cu un nivel scăzut de hipertermie. Ei pot prezenta stare de prostrație, mișcări ezitante sau înțepenite, însă cel mai evident semn al PDNS este prezența unor macule și papule cutanate, de formă neregulată, culoare roșie – purpurie în special la nivelul membrelor
posterioare și în zona perineală, rareori distribuite generalizat. După aproximativ 2-3 săptămâni, aceste leziuni sunt acoperite de cruste de culoare închisă. Leziunile se estompează treptat, lăsând uneori cicatrici.
c. Sindromul reproducător porcin (PRD)
Este caracterizat prin avorturi în ultima perioadă a gestației sau prin nașterea de purcei morți, cu sau fără prezența altor agenți patogeni abortigeni cunoscuți.
Aceste semne par a fi semnul distinctiv al infecției cu PCV-2 la scroafe. Leziuni anatomopatologice
a. Sindromul de epuizare multisistemică post-înțărcare Macroscopic, limfonodurile sunt mult mărite în volum și palide pe suprafața de secțiune, urmând ca acestea să revină la mărimea normală, sau chiar să sufere atrofie în fazele avansate ale PMWS; corticala timusului este atrofiată, iar tonsilele sunt reduse în volum. La unii indivizi se pot observa infarcte splenice.
Leziunile pulmonare sunt frecvent întâlnite, iar severitatea acestora este în strânsă legătură cu durata de evoluție a bolii și cu prezența infecțiilor concurente. Pulmonii sunt măriți în volum, necolabați, având o consistență elastică și un aspect marmorat. Unii lobuli sunt atelectazici. Uneori poate fi afectat și ficatul, acesta fiind redus în volum, palid și ferm, cu suprafața fin granulară, atunci când indivizii manifestă icter.
.png)
Țesutul conjunctiv interlobular poate fi proeminent. Rinichii pot fi măriți în volum cu prezența unor focare de culoare albă pe suprafața cortica lei. Unii indivizii afectați pot prezenta ulcere gastrice și miocardită multifocală limfohistiocitară.
Leziunile histopatologice caracteristice la nivelul limfonodurilor sunt depleția limfoidă, infiltratul cu histiocite mari și celule gigant multinucleate. Se pot observa incluzii virale intracitoplasmatice în histiocite și celule dendritice. Apar leziuni variabile, de la pneumonie interstițială limfohistiocitară, până la bronhopneumonie interstițială granulomatoasă cu bronșiolită, fibroză peribronșiolară și bronșiolită fibrinoasă, în cazurile severe.
La nivelul ficatului, leziunile variază de la apoptoza cu infiltrat limfocitar în zonele portale, până la hepatită periportală limfohistiocitară extensivă cu necroza difuză a hepatocitelor. Pot fi observate leziuni de nefrită interstițială limfohistiocitară difuză severă.
b. Sindromul dermatită-nefropatie PDNS este ușor de identificat datorită leziunilor de la nivel cutanat, care apar, în funcție de stadiul evolutiv, ca macule, papule sau cruste, rotunde sau neregulate, denivelate, de culoare roșie-purpurie cu centrul negru. Indivizii care mor subit prezintă rinichii măriți în volum, palizi, corticala prezentând un aspect fin granular și leziuni miliare de culoare roșietică, precum și edem al bazinetului renal. În forma cu o evoluție mai lungă se observă glomerulonefrită cronică. În mod normal, atât leziunile cutanate, cât și cele renale sunt prezente în PDNS, dar la unii indivizi, leziunile cutanate sau renale pot apărea singure. Limfonodurile pot fi mărite și de culoare roșie. Infarctele splenice pot fi, de asemenea, prezente.
Histologic, maculele și papulele, sunt observate ca leziuni cutanate hemoragice și necrotice, asociate cu vasculită necrotică.
La nivel renal se identifică leziuni de glomerulită hemoragică în stadiile incipiente, apoi formarea de hialin în spațiul Bowman, nefrită interstițială nesupurată și chistizarea tubilor uriniferi. Se evidențiază leziuni de miocardită fibrino-necrotică.
La nivelul limfonodurilor, leziunile întâlnite sunt similare celor din PMWS.
c. Sindrom reproducător suin Purceii născuți morți și neviabili prezintă congestie hepatică pasivă cronică și hipertrofie cardiacă cu zone multifocale de decolorare a miocardului. Leziunea microscopică corespunde unei miocardite nesupurate, fibrinoasă și/sau necrotică.
Diagnosticul de laborator Pentru toate cele trei sindroame, cele mai utilizate metode pentru stabilirea unui dignostic de certitudine sunt: PCR, imunohistochimia și hibridizarea in situ. Probele ce trebuie trimise spre laboratoarele de diagnostic sunt: organe cu leziuni, biopsii cutanate
din zonele ce prezintă leziuni sau avortoni.
Sindromul reproducător și respirator suin (PRRS)
Etiologie
Virusul sindromului reproducător și respirator suin este un virus ARN monocatenar, capsidat, cu dimensiuni cuprinse între 45 și 80 nm, ce face parte din genul Arterivirus, clasificat în familia Arteriviridae.
Epidemiologie
Virusul PRRS este prezent în majoritatea regiunilor din lume unde se cresc porci. La animalele infectate, virusul este prezent în salivă, secreții nazale, urină, spermă, și uneori în fecale. Femelele gestante infectate în ultima perioadă a gestației vor elimina virusul prin lapte. Suinele sunt susceptibile la contaminare pe mai multe căi: intranazal, intramuscular, intrauterin sau vaginal. Unele studii arată faptul că leziunile cutanate sunt cea mai frecventă cale de contaminare, în timpul comportamentelor agresive între indivizi, virusul eliminat prin salivă, poate ajunge cu
ușurință la nivelul țesutului cutanat al altui individ. PRRSV se transmite, de asemenea, și pe cale verticală, transplacentar, ducând la moarte embrionară sau fetală. Virusul este capabil să traverseze bariera placentară în al treilea și, probabil, în al doilea trimestru al gestației. Astfel,
purceii se pot naște viremici și transmit virusul timp de 112 zile după infecție.
.png)
Patogeneză
În urma expunerii la virus, acesta inițiază replicarea la nivelul macrofagelor locale, urmând ca apoi să se răspândească rapid, în special la nivelul organelor limfoide și plămânilor. Viremia apare la 12-24 de ore post infecție. Infecția cu virusul PRRS are ca rezultat distrugerea macrofagelor alveolare mature, ceea ce a dus la ipoteza că infecția are ca rezultat suprimarea imunității. Replicarea acestuia în macrofagele de la nivelul pulmonilor și a țesutului limfoid induce leziuni și semne clinice prin mecanisme diverse: apoptoza celulelor infectate, secreția
citokinelor inflamatorii, activarea celulelor B policlonale și reducerea fagocitozei bacteriene datorată morții macrofagelor, ceea ce crește riscul apariției septicemiei.
Tablou clinic
Boala este caracterizată prin avorturi în ultima perioadă a gestației, avorturi la termen și tulburări respiratorii. Expresia clinică a PRRS variază de la asimptomatic până la sever, fiind influențată de varietatea virusului, de imunitatea și susceptibilitatea gazdei, precum și de infecțiile concurente.
Prima fază a formei epidemice a PRRS este caracterizată de anorexie și letargie la 5-75% din efectiv, cu o evoluție de 3-7 zile. Indivizii din creșă și tineretul sunt cei mai afectați din punct de vedere clinic, prezentând hipertermie (39-41ºC), letargie, stare de stupoare datorită infecției sistemice,
urmate de dispnee, în special în timpul expirației. Vârsta la care indivizii sunt cei mai susceptibili pentru boala respiratorie este cea de 4-10 săptămâni. Scroafele ajunse la maturitatea sexuală și vierii pot prezenta anorexie, hipertermie, letargie, insuficiență respiratorie și emeză. La
scroafe apare hiperemie și cianoză cutanată, auriculară (boala urechilor albastre), abdominală, vulvară și la nivelul râtului.
A doua fază include insuficiență repro- ductivă la scroafele cu viremie în al 3-lea trimestru de gestație, ceea ce duce la creșterea numărului de purcei fătați morți, eliminare de fetuși mumifiați, fătări premature și purcei debili. Rata de apariție a fetușilor mumifiați și a celor născuți morți
este de 25-35%, pe când rata avorturilor trece ușor peste 10%. Scroafele aflate în perioada de lactație sunt anorexice și suferă de agalaxie, ceea ce duce la o mortalitate a purceilor de 30%-50% înainte de înțărcare.
Purceii sugari dezvoltă o insuficiență respiratorie cu un tablou clinic caracteristic ce constă în dificultate în respirație, respirație abdominală, cu gura deschisă. Focarele cu forma reproductivă durează între 1 și 4 luni conform studiilor, în funcție de statusul de sănătate inițial al indivizilor
și de facilitățile de bunăstare a fermei. În schimb, forma respiratorie poate evolua cronic, reducând sporul zilnic de creștere cu 85% și crescând mortalitatea la 10-25%.
Leziuni anatomopatologice
Macroscopic, leziunile pulmonare variază de la modificări ușoare ce sunt caracterizate prin necolabarea pulmonilor la deschiderea toracelui și păstrarea amprentelor costale, până la modificări severe caracterizate prin consolidare pulmonară în caz de suprainfecție bacteriană și hipertrofie a
limfonodurilor traheobronşice și mediastinale. Pneumonia interstițială poate fi lobulară sau difuză. Pulmonii apar cu un aspect marmorat.
În cazul formei reproductive, leziunile caracteristice sunt hemoragii segmentare sau difuze la nivelul cordonului ombilical și arterită ombilicală necrozantă. La unii avortoni apare ascită, hidrotorax, edem la nivel perirenal, mezenteric și al ligamentului splenic.
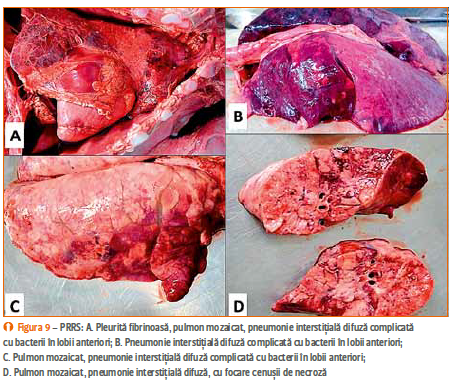
Purceii fătați morți și cei neviabili prezintă congestie și edem pulmonar, pneumonie interstițială necrotică, dilatație cardiacă și transsudate în marile cavități. Uneori au fost observate și hemoragii cutanate, edem al pleoapelor și al regiunii periorbitale.
La purceii cu vârste cuprinse între 1 și 3 luni apar: pneumonie interstițială, bronhopneumonie purulentă în special la nivelul lobilor anteriori; consolidări lobulare în lobii diafragmatici, iar limfonodurile apar mărite în volum. Scroafele infectate cu PRRS au leziuni pulmonare și sistemice tipice,
endometrită, miometrită și placentită. Histologic, în forma respiratorie, modi- ficările relevante sunt de pneumonie inter- stițială caracterizată prin îngroșarea pereților alveolari, datorată infiltrării macrofagelor și limfocitelor, precum și hiperplazia pneumocitelor de tip II. La nivelul spațiilor
alveolare se pot identifica detritusuri celulare.
Se observă necroze limfoide, hiperplazie limfoidă nodulară în organele limfoide; miocardită limfoplasmocitară și encefalită nesupurată. În forma reproductivă se observă arterită segmentară la nivelul ombilicului, pulmonilor, cordului și rinichilor. Agregatele de limfocite, plasmocite și macrofage sunt prezente în vasele coronariene, venele porte și la nivelul substanței albe din cerebel. Se observă edem și infiltrat cu limfocite și histiocite în interstițiul uterin și în jurul vaselor uterine, precum și hiperplazia centrilor germinativi limfonodali cu celule multinucleate.
Diagnosticul de laborator
Diagnosticul de certitudine se face prin examen PCR, hibridizare in situ, imunohistochimic sau prin imunofluorescență directă. Se recomandă recoltarea următoarelor probe: pulmon, tonsile, limfonoduri, cord, creier, timus, splină, rinichi, avorton în formaldehidă 10% pentru
IHC sau la gheață pentru PCR și IFD.
Bibliografie
1. Allan G.M., Ellis J.A., (2000), Porcine Circoviruses: A Review. J Vet Diagn Invest 12: 3-14;
2. Allan,G.M., Kennedy,S., McNeilly,F., Foster,J.C., Ellis,J.A., Krakowka,S.J., Meehan,B.M., Adair,B.M., (1999). Experimental reproduction of severe wasting disease by co-infection of pigs with porcine circovirus and porcine parvovirus. J.Comp.Pathol.121: 1–11.
3. Andraud, M., Grasland, B., Durand, B., Cariolet, R., Jestin, A., Madec, F., Pierre, J.S., Rose, N., (2009a). Modelling the time-dependent transmission rate for porcine circovirus type 2 (PCV2) in pigs using data from serial transmission experiments. J. R. Soc. Interface 6: 39–50.
4. Andraud, M., Rose, N., Grasland, B., Pierre, J.S., Jestin, A., Madec, F., (2009b). Influence of husbandry and control measures on porcine circovirus type 2 (PCV- 2) dynamics within a farrow-to-finish pig farm: a modelling approach. Prev. Vet. Med. 92: 38–51.
5. Becher, P., Orlich, M., Kosmidou, A., Konig, M., Baroth, M., Thiel, H.J., (1999). Genetic diversity of pestivirus: identification of novel groups and implications for classification. Virology 262: 64–71.
6. Bierk M.D., Dee S.A., Rossow K.D., Otake S., Collins J.E., Molitor T.W., (2001). Transmission of porcine reproductive and respiratory syndrome virus from persistently infected sows to contact controls. Can J Vet Res 65[4]: 261-266
7. Blaha, Th., (1992). Epidemiological investigations into PEARS in Germany: consequences in fattening pigs. Proceedings of 12th International Pig Veterinary Society Congress, The Hague, The Netherlands, 17–20 August, 126
8. Blome, S., Staubach, C., Henke, J., Carlson, J., Beer, M., (2017). Classical Swine Fever—An Updated Review. Viruses 9(4): 86-128
9. Bool, P.H., Ordas, A., Sanchez Botija, C., (1969). El diagnostico de la peste porcina africana por inmunofluorescencia (The diagnosis of African swine fever by immunofluorescence). Bulletin of the OIE 72: 819–839
10. Carasova,P., Celer,V., Takacova,K., Trundova,M., Molinkova,D., Lobova,D., (2007). The levels of PCV2 specific antibodies and viremia in pigs. Research in Veterinary Science 83[2]: 274–278.
11. Carlsson U., Wallgren P., Renstrom L.H.M., Lindberg A., Eriksson H., Eliasson- Selling L., Lundeheim N., Norregard E., Thorn C., Elvander M., (2009). Emergence of Porcine Reproductive and Respiratory Syndrome in Sweden: Detection, Response and Eradication. Transbondary and Emerging Diseases 59: 121-131
12. Carrasco L., Chacón-M de Lara F., Martín de las Mulas J., Gómez-Villamandos Jc., Hervás J.,(1996). Virus association with lymphocytes in acute African swine fever. Veterinary Research, BioMed Central 27[3]: 305-312
13. Cătoi C., Diagnostic Necropsic Veterinar, Ed. Academicpres, Cluj-Napoca, 2003;
14. Clark E.G., (1997). Post-weaning multisystemic wasting syndrome. Proceed-ings of the American Association of Swine Practitioners 499-501
15. Colgrove G.S., Haelterman E.O., Coggins L., (1969). Pathogenesis of African swine fever in young pigs. Amer J Vet Res 1343:1359
16. Darwich L., Segales J., Mateu E., (2004). Pathogenesis of postweaning multisystemic wasting syndrome caused by Porcine circovirus 2: an immune riddle. Arch Virol 149: 857-874
17. De Tray (1957). African Swine Fever: a review. Bull. Dis. Afr., 5: 475–478
18. Dixon L.K., Chapman D., Abrams C., Rowlands R. (2011) Asfivirus. In: Tidona C., Darai G. (eds) The Springer Index of Viruses. Springer, New York, NY
19. Drolet, R., Thibault, S., D’Allaire, S., Thompson, J., Done, S., (1999). Porcine dermatitis and nephropathy syndrome (PDNS): an overview of the disease. Swine Health Prod. 7 (6): 283–285
20. Durand, S.V., Hulst, M.M., (2009). Activation and modulation of antiviral and apoptotic genes in pigs infected with classical swine fever viruses of high, moderate or low virulence. Arch. Virol. 154 (9): 1417-1431.
21. Edwards, S., (2000). Survival and inactivation of classical swine fever virus. Vet. Microbiol. 73: 175–181.
22. Floegel-Niesmann, G., Blome, S., Gerβ-Dulmer, H., Bunzenthal, C., Moennig, V., (2009). Virulence of classical swine fever virus isolates from Europe and other areas during 1996 until 2007. Vet. Microbiol. 139, 165–169.
23. Gomez-Villamandos J.C., Hervas J., Mendez A., Carrasco L., de la Mulas J.M., Villeda C.J., Wilkinson P.J., Sierra M.A., (1995). Experimental African swine fever: apoptosis of lymphocytes and virus replication in other cells. Journal of General Virology 76: 2399-2405
24. Grau-Roma L., Hjulsager C.K., Sibila M., Kristensen C.S., López-Soria S., Enoe C., (2009). Infection, excretion and seroconversion dynamics of porcine circovirus type 2(PCV2) in pigs from post-weaning multisystemic wasting syndrome (PMWS)affected farms in Spain and Denmark. Vet Microbiol 135: 272–82
25. He, Y.X., Hua, R.H., Zhou, Y.J., Qiu, H.J., Tong, G.Z., (2007). Interference of porcine reproductive and respiratory syndrome virus replication on MARC145 cells using DNA-based short interfering RNAs. Antiviral Res. 74: 83– 91.
26. Jamin A., Gorin S., Cariolet R., Le Potier M.F., Kuntz-Simon G., (2008). Classical swine fever virus induces activation of plasmacytoid and conventional dendritic cells in tonsil, blood, and spleen of infected pigs. Vet Res 39: 1-17
27. Jamnikar Ciglenecki, U., Grom, J., Toplak, I., Jemersic, L., Barlic-Maganja, D., (2008).Real-time RT-PCR Assay for rapid and specific detection of classical swine fevervirus: comparison of SYBR green and TaqMan MGB detection methods usingnovel MGB probes. J. Virol. Methods 147: 257–264
28. King, D.P., Reid, S.M., Hutchings, G.H., Grierson, S.S., Wilkinson, P.J., Dixon, L.K., Bastos, A.D., Drew, T.W., (2003). Development of a TaqMan PCR assay with internal amplification control for the detection of African swine fever virus. J. Virol. Methods 107: 53–61.
29. MacLachlan N. J., Dubovi E.J.,(2011). Fenner’s Veterinary Virology 4th edition. Editor: Academic Press, an imprint of Elsevier, 32 Jamestown Road, London NW1 7BY, UK
30. Malmquist W.A., Hay D., (1960) Hemadsorption and cytopathic effect produced by African swine fever virus in swine bone marroe and buffy coat cultures. American Journal of Veterinary Research 21: 104-108
31. Mebus C.A., McVicar J.W., Dardiri A.H., (1983). Comparison of the pathology of high and low virulence African swine fever virus infections. In: CEC/FAO Expert Consultation in Africa Swine fever Research Seminar, Sardinia
32. Minguez I., Rueda A., Dominguez J., Sanchez-Vizcaino J.M., (1998). Double Labeling Immunohistological Study of African Swine Fever Virus-infected Spleen and Lymph Nodes. Vet. Pathol 25: 193-198
33. Oleksiewicz, M.B., Botner, A., Madsen, K.G., Storgaard, T., (1998). Sensitive detection and typing of porcine reproductive and respiratory syndrome virus by RT-PCR amplification of whole viral genes. Vet. Microbiol. 64 (1): 7–22.
34. Opriessing T., Mckeown N.E., Harmon K.L., Meng X.J., Halbur P.G., (2006). Porcine circovirus type 2 infection decreases the efficacy of a modified live porcinereproductive and respiratory syndrome virus vaccine. Clin Vaccine Immunol 13(8): 923-929
35. Opriessnig, T., Patterson, A. R., Jones, D. E., Juhan, N. M., Meng, X-J. and Halbur, P. G. (2009): Limited susceptibility of three different mouse (Mus musculus) lines to porcine circovirus2 infection and associated lesions. Can. J. Vet. Res. 73, 81–85.
36. Park J.S., Ha Y., Kwon B., Cho K.D., Lee B.H., Chae C., (2009). Porcine epidemic diarrhea virus infects and replicates in porcine alveolar macrophages. Virus Research, 191: 143-152.
37. Rosell C., Segales J., Ramos-Vara J.A., Folch J.M., Rodriguez-Arrioja G.M., Duran C.O., Balach M., Plana-Duran J., Domingo M., (2000). Identification of porcine circovirus in tissues of pigs with porcine dermatitis and nephropathy. Veterinary Record 146: 40-43
38. Sanchez Botija C., (1982). African swine fever.New developments. Rev Scitech Off int Epiz 1(4): 1065-1094
39. Segalés, J., Piella, J., Marco, E., Mateu-de-Antonio, E.M., Espuña, E., Domingo, M., (1998). Porcine dermatitis and nephropathy syndrome in Spain. Vet. Rec. 142: 483–486.
40. Segalés J., Rosell C., Domingo M., (2004). Pathological findings associated with naturallyacquired Porcine Circovirus Type 2 associated disease. Vet Microbiol 98:137–49
41. Segales J., Allan G.M., Domingo M., (2005), Porcine circovirus diseases. Animal Health Research Reviews 6[2]: 119-142;
42. Susa, M., Konig, M., Saalmuller, A., Reddehase, M.J., Thiel, H.J., 1992. Pathogenesis of classical swine fever: B-lymphocyte deficiency caused by hog cholera virus. J. Virol. 66: 1171-1175.
43. Wagstrom E.A., Chang C.C., Yoon K.J., Zimmerman J.J., (2001). Shedding of porcine reproductive and respiratory syndrome virus in mammary gland serections of sows. AJVR 62[12]: 1876-1880
44. Weesendorp, E., Backer, J., Stegeman, A., Loeffen, W., (2011). Transmission of classical swine fever virus depends on the clinical course of infection which is associated with high and low levels of virus excretion. Vet. Microbiol. 147: 262–273.
45. Wilkinson,P.J., Wardley,R.C., (1978). The replication of African swine fever virus in pig endothelial cells. British Veterinary Journal 134(3): 280–282
46. Wills, R.W., Zimmerman, J.J., Yoon, K.J., Swenson, S.L., McGinley, M.J., Hill, H.T., Platt, K.B., Christopher-Hennings, J., Nelson, E.A., (1997). Porcine reproductive and respiratory syndrome virus: a persistent infection. Vet. Microbiol. 55: 231–240.
47. Yoon K.J., Zimmerman J.J., Chang C.C., Cancel-Tirado S., Harmon K.M., Mcginley M.J., (1999). Effect of challenge dose and route on porcine reproductive
and respiratory syndrome virus (PRRSV) infection in young swine. Veterinary Research 30[6]: 629-638
48. Zachary J.F., (2016). Pathologic basis of veterinary disease 6th edition. Editor:Elsevier, 3251 Riverport Lane, St. Louis, Missouri 63043
49. Zimmerman J.J., Karriker L.A., Ramirez A., Schwartz K.J., Stevenson G.W.,(2012). Diseases of swine 10th edition. Editor: John Wiley & Sons Ltd, The Atrium, Southern Gate, Chichester, West Susses, PO19 8SQ, UK
May
10
Această lucrare sintetizează caracteristicile histostructurale ale intestinului, ficatului și vezicii biliare, subliniind adaptările care permit digestia eficientă, absorbția nutrienților și apărarea imună. Intestinul subțire prezintă vilozități lungi, glande Lieberkühn profunde și plăci Peyer extinse (Young et al., 2020). Ficatul se remarcă prin lobuli bine conturați, triade portale proeminente și hepatocite active metabolic, iar vezica biliară se caracterizează prin pliuri mucoase care permit stocarea și evacuarea bilei (Trautmann & Fiebiger, 2015). Aceste caracteristici le permit suinelor să digere eficient o dietă variată, cu o detoxificare hepatică eficientă şi un sistem imunitar robust. Informațiile prezentate sunt esențiale pentru interpretarea modificărilor fiziologice și patologice induse de factori nutriționali, toxici sau microbieni și oferă un cadru pentru cercetare aplicată în medicină veterinară și nutriție animală (Moeser, 2007).
Mar
12
Leucoza enzootică bovină (LEB) reprezintă o problemă economică multilaterală şi complexă, cu implicaţii semnificative atât la nivelul abatoarelor, cât și al întregii producţii zootehnice. Impactul său financiar se manifestă prin pierderi directe, indirecte și de oportunitate. Etiologia leucozei bovine este reprezentată de un retrovirus oncogen exogen, de tip C, din grupul virusului T-limfotrop uman (HTLV) și al virusului leucozei bovine (BLV). În pofida instalării unei infecţii persistente, majoritatea pacienţilor rămân asimptomatici, însă un număr mic de indivizi dezvoltă limfosarcom, ceea ce conduce la identificarea tumorilor în timpul inspecţiei post-mortem din abatoare. Manifestările clinice sunt direct corelate cu localizarea și invazia neoplazică a sistemelor de organe.
Aug
18
Pomeranianul este o rasă canină de talie mică, remarcabilă prin aspectul său distinctiv și comportamentul alert, inteligent și afectuos. Deși foarte apreciat ca animal de companie, acest câine este predispus la multiple afecțiuni asociate morfologiei sale miniaturale și particularităților genetice. Sunt descrise punctual cele mai frecvente patologii întâlnite la această rasă, incluzând afecțiuni dentare (gingivită, parodontită, anomalii de ocluzie), fontanela persistentă și hidrocefalia congenitală, hipoglicemia neonatală, alopecia X, strănutul invers, colapsul traheal, cardiomiopatiile (congenitale și dobândite) și luxația patelară, cu particularitățile fiziopatologice ale rasei. Managementul eficient al acestor afecțiuni presupune o abordare preventivă, monitorizare riguroasă și, în cazurile severe, intervenții chirurgicale sau tratamente farmacologice orientate pe vulnerabilitățile acestei rase, cunoașterea acestora devenind esențială pentru menținerea unei bune calități a vieții și prevenirea complicațiilor sistemice.
Apr
16
Sănătatea animalelor de fermă – o provocare permanentă pentru medicii veterinari
Inscriete la noutatile noastre